Bài viết Cúm 1918: Đại dịch lớn nhất lịch sử được dịch bởi Bác sĩ Nguyễn Thành Luân – ICU bệnh viện Hoàn Mỹ Cửu Long và Bác sĩ đa khoa Nguyễn Mai Anh – Đại học Y Dược Hải Phòng từ bài viết 1918 Influenza: the Mother of All Pandemics.
Tác giả: Jeffery K. Taubenberger * và David M. Morens †.
* Tiến sĩ Taubenberger là chủ tịch của Khoa Bệnh học Phân tử tại Viện Bệnh học Lực lượng Vũ trang, Rockville, Maryland. Các nghiên cứu của ông bao gồm sinh bệnh học phân tử và sự phát triển của các chủng virus cúm. † Tiến sĩ Morens là một nhà dịch tễ học có mối quan tâm lâu dài về các bệnh truyền nhiễm mới nổi, virus học, y học nhiệt đới và lịch sử y khoa. Từ năm 1999, ông làm việc tại Viện Dị ứng và Bệnh Truyền nhiễm Quốc gia.
Giới thiệu
Đại dịch cúm “Tây Ban Nha” năm 1918-1919, gây ra khoảng 50 triệu trường hợp tử vong trên toàn thế giới, vẫn là một cảnh báo đáng ngại cho sức khỏe cộng đồng. Nhiều câu hỏi về nguồn gốc, các đặc điểm dịch tễ bất thường và những vấn đề bệnh học cơ bản của nó vẫn chưa được trả lời. Do đó, ý nghĩa đối với sức khỏe cộng đồng của đại dịch vẫn còn nghi ngờ ngay cả khi chúng ta đang vật lộn với sự xuất hiện đáng sợ của một đại dịch do H5N1 hoặc virus khác gây ra. Gần đây có thông tin mới về virus 1918, ví dụ, giải trình tự toàn bộ bộ gen từ các mô tử thiết lưu trữ; nhưng, bộ gen virus không thể cung cấp câu trả lời cho một số câu hỏi quan trọng. Hiểu về đại dịch 1918 và ý nghĩa của nó đối với các đại dịch trong tương lai đòi hỏi phải thực nghiệm cẩn thận và phân tích lịch sử sâu rộng.
Ước tính một phần ba dân số thế giới (hoặc khoảng 500 triệu người) đã bị nhiễm và có biểu hiện bệnh rõ ràng trên lâm sàng [1,2] trong đại dịch cúm 1918-1919. Bệnh tình đặc biệt nghiêm trọng. Tỷ lệ tử vong trong số các ca nhiễm là >2.5%, so với <0.1% trong các đại dịch cúm khác [3,4]. Tổng số người chết được ước tính là khoảng 50 triệu [5-7] và được cho là lên tới 100 triệu [7].
Tác động của đại dịch này không chỉ giới hạn ở 1918-1919. Tất cả các đại dịch cúm A kể từ thời điểm đó, và thực tế là hầu hết các trường hợp mắc cúm A trên toàn thế giới (ngoại trừ nhiễm cúm ở người do virus cúm gia cầm như H5N1 và H7N7), đã được gây ra bởi hậu duệ của virus 1918, bao gồm cả virus cúm H1N1 “đột biến” và virus H2N2 và H3N2 tái tổ hợp. Virus H2N2 và H3N2 tái tổ hợp bao gồm các gen chủ chốt của virus 1918, được cập nhật bởi các gen cúm gia cầm hợp nhất sau đó, chúng mã hóa cho các protein bề mặt mới, làm cho virus 1918 thực sự là “mẹ” của tất cả các đại dịch.
Năm 1918, nguyên nhân của bệnh cúm ở người và mối liên hệ của nó với cúm gia cầm và cúm lợn vẫn chưa được biết rõ. Mặc dù có sự tương đồng về mặt lâm sàng và dịch tễ học với đại dịch cúm năm 1889, 1847 và thậm chí trước đó nữa, nhưng nhiều người đã đặt câu hỏi liệu một căn bệnh gây tử vong bùng nổ như vậy có thể là cúm hay không. Câu hỏi đó đã không bắt đầu được giải quyết cho đến những năm 1930, khi các virus cúm có liên quan chặt chẽ (hiện được gọi là virus H1N1) được phân lập, đầu tiên là từ lợn và ngay sau đó là từ người. Các nghiên cứu dịch tễ học đã sớm liên kết cả hai loại virus này với đại dịch năm 1918 [8]. Các nghiên cứu tiếp theo chỉ ra rằng hậu duệ của virus 1918 vẫn còn gây dịch bệnh kéo dài ở lợn. Chúng có lẽ cũng tiếp tục lưu hành ở người, trải qua sự trôi dạt kháng nguyên dần dần và gây ra dịch bệnh hàng năm, cho đến những năm 1950. Với sự xuất hiện của một đại dịch H2N2 mới vào năm 1957 (“cúm Châu Á”), hậu duệ trực tiếp của virus cúm H1N1 trong đại dịch 1918 đã biến mất hoàn toàn khỏi con người, mặc dù dòng dõi có liên quan vẫn gây dịch bệnh kéo dài khu trú ở lợn. Nhưng vào năm 1977, virus cúm người đột nhiên xuất hiện trở lại từ một tủ đông trong phòng thí nghiệm [9]. Chúng tiếp tục lưu hành tại chổ và gây dịch bệnh.
Do đó, vào năm 2006, 2 dòng dõi hậu duệ chính của virus H1N1 1918, cũng như 2 dòng tái tổ hợp khác, vẫn tồn tại trong tự nhiên: một dòng dịch cúm/bệnh cúm H1N1 ở người, một dòng cúm H1N1 gây bệnh ở lợn (còn được gọi là cúm lợn cổ điển), và dòng virus H3N2 tái tổ hợp ở người – giống như virus H1N1 ở người, đã dẫn đến một dòng H3N2 ở lợn. Tuy nhiên, không có thế hệ hậu duệ virus nào tiếp cận được khả năng gây bệnh của virus mẹ năm 1918. Rõ ràng, các dòng H1N1 và H3N2 lợn không thường lây nhiễm ở người, và cả hai dòng H1N1 và H3N2 ở người đều có liên quan đến tỷ lệ mắc bệnh và tử vong thấp hơn so với virus 1918. Trên thực tế, tỷ lệ tử vong hiện tại của H1N1 thậm chí còn thấp hơn so với các chủng thuộc dòng H3N2 (phổ biến từ năm 1968 đến nay). Virus H1N1 xuất phát từ chủng 1918, cũng như virus H3N2, hiện đã đồng thời lưu hành trên toàn thế giới trong 29 năm và cho thấy rất ít bằng chứng về sự tuyệt chủng sắp xảy ra.
Cố gắng hiểu những gì đã xảy ra
Đến đầu những năm 1990, 75 năm nghiên cứu đã không trả lời được câu hỏi cơ bản nhất về đại dịch 1918: tại sao nó lại gây tử vong như vậy? Không có virus từ chủng 1918 đã được phân lập, nhưng tất cả các hậu duệ rõ ràng của nó gây ra bệnh nhẹ hơn đáng kể ở người. Hơn nữa, việc kiểm tra dữ liệu về tỷ lệ tử vong từ những năm 1920 cho thấy rằng trong vòng một vài năm sau 1918, dịch cúm đã biến thành một mô hình dịch bệnh hàng năm liên quan đến tình trạng trôi dạt các chủng và giảm đáng kể tỷ lệ tử vong. Có phải một số sự kiện di truyền virus quan trọng đã tạo ra virus 1918 có khả năng gây bệnh nổi bật và rồi một sự kiện di truyền quan trọng khác xảy ra ngay sau đại dịch 1918 để tạo ra một loại virus H1N1 bị suy yếu?
Năm 1995, một nhóm khoa học đã tìm thấy các tài liệu tử thiết cúm lưu trữ được thu thập vào mùa thu năm 1918 và bắt đầu giải trình tự các đoạn RNA nhỏ của virus để xác định cấu trúc gen của virus cúm gây bệnh [10]. Nhờ những nỗ lực này mà hiện nay người ta xác định được trình tự bộ gen hoàn chỉnh của 1 loài virus và một phần trình tự của 4 loại virus khác. Dữ liệu ban đầu từ các nghiên cứu trên [11-17] và một số bài tổng quan bao trùm các khía cạnh khác nhau của đại dịch 1918 đã được công bố gần đây [18-20] và xác nhận rằng virus 1918 có khả năng là tổ tiên của cả 4 dòng H1N1 và H3N2 của lợn và người, cũng như dòng dõi H2N2 đã “tuyệt chủng”. Không có đột biến nào được biết có liên quan đến khả năng gây bệnh cao ở các loại virus cúm người hoặc động vật khác đã được tìm thấy trong bộ gen 1918, nhưng các nghiên cứu đang tiến hành để xác định các yếu tố độc lực đang cho kết quả thú vị. Tuy nhiên, dữ liệu trình tự 1918 để lại những câu hỏi chưa được trả lời về nguồn gốc của virus [19] và về dịch tễ học của đại dịch.
Đại dịch cúm 1918 xảy ra khi nào và ở đâu?
Trước và sau 1918, hầu hết các đại dịch cúm đã phát triển ở Châu Á và lan rộng từ đó đến phần còn lại của thế giới. Sự phân định rõ ràng về điểm xuất phát địa lý, đại dịch 1918 lan truyền đồng thời trên dưới 3 đợt khác nhau trong khoảng thời gian ̴12 tháng vào năm 1918-1919, ở Châu Âu, Châu Á và Bắc Mỹ (đợt đầu tiên được mô tả rõ nhất ở Hoa Kỳ vào tháng 3 năm 1918). Dữ liệu lịch sử và dịch tễ học không đủ để xác định nguồn gốc địa lý của virus [21] và phân tích phát sinh loài gần đây của bộ gen virus 1918 không đặt virus vào bất kỳ bối cảnh địa lý nào [19].
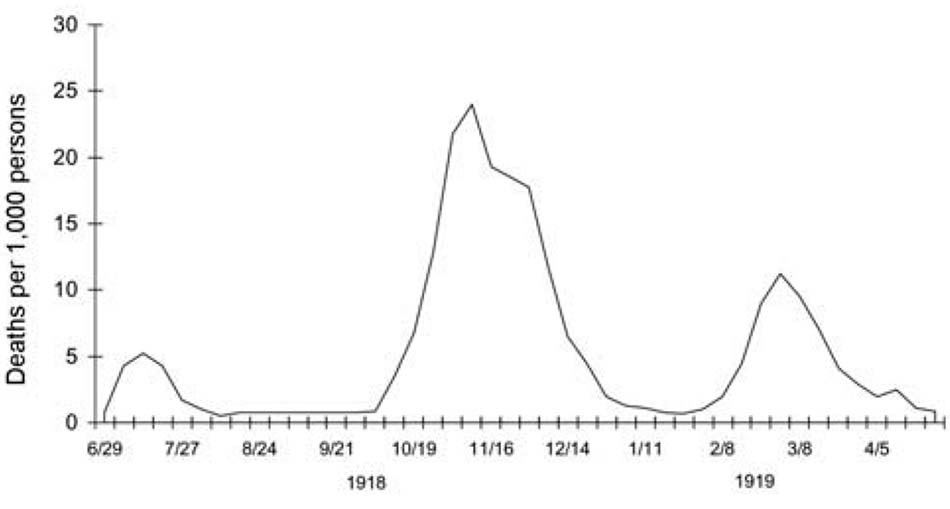
Mặc dù vào năm 1918, cúm không phải là bệnh có thể báo cáo trên toàn quốc và các tiêu chuẩn chẩn đoán về cúm và viêm phổi là mơ hồ, nhưng tỷ lệ tử vong do cúm và viêm phổi ở Hoa Kỳ đã tăng mạnh vào năm 1915 và 1916 do dịch bệnh hô hấp lớn bắt đầu vào tháng 12 năm 1915 [22]. Tỷ lệ tử vong sau đó giảm nhẹ vào năm 1917. Đợt đại dịch cúm đầu tiên xuất hiện vào mùa xuân năm 1918, sau đó nhanh chóng xảy ra các đợt dịch thứ hai và thứ ba gây tử vong nhiều hơn tương ứng vào mùa thu và mùa đông năm 1918-1919 (Hình 1). Liệu có phải virus H1N1 thích nghi kém đã bắt đầu lây lan vào năm 1915, gây ra một số bệnh cảnh nặng nhưng chưa đủ sức để bắt đầu một đại dịch? Dữ liệu đồng nhất khả năng này đã được báo cáo cùng thời điểm từ các khu quân đội Châu Âu [23], nhưng một lập luận phản đối là nếu một chủng với hemagglutinin (HA) mới gây ra bệnh cảnh đủ ảnh hưởng đến tỷ lệ tử vong toàn Hoa Kỳ do viêm phổi và cúm thì nó đã gây ra một đại dịch sớm hơn, và cuối cùng, khi đại dịch xảy ra vào năm 1918, nhiều người sẽ có miễn dịch hoặc ít nhất là được bảo vệ miễn dịch một phần. Các sự kiện “báo trước” vào năm 1915, 1916 và thậm chí vào đầu năm 1918, nếu chúng xảy ra, sẽ rất khó để xác định.
Có phải 3 đợt cúm vào 1918-1919 được gây ra bởi cùng một loại virus?
Nếu vậy, làm thế nào và tại sao chúng xảy ra?
Các ghi chép lịch sử từ thế kỷ 16 cho thấy những đại dịch cúm mới có thể xuất hiện bất cứ lúc nào trong năm, không nhất thiết phải theo mô hình mùa đông hàng năm quen thuộc của những năm giữa các đại dịch, có lẽ là do các virus cúm mới thay đổi thích nghi một cách khác đi khi chúng tìm thấy một quần thể người rộng lớn hoặc rất dễ mắc bệnh. Sau đó, vì phải đối mặt với áp lực chọn lọc của miễn dịch quần thể, những virus gây đại dịch này bắt đầu trôi dạt về mặt di truyền và cuối cùng biến thành một mô hình tái phát dịch bệnh hàng năm do các biến thể virus trôi dạt.
Trong đại dịch 1918-1919, đợt dịch đầu tiên hoặc mùa xuân bắt đầu vào tháng 3 năm 1918 và lan rộng không đều qua Hoa Kỳ, Châu Âu và có thể cả Châu Á trong 6 tháng tiếp đó (Hình 1). Tỷ lệ mắc bệnh cao, nhưng tỷ lệ tử vong ở hầu hết các nơi không được đánh giá là cao hơn mức bình thường. Đợt dịch thứ hai hoặc mùa thu lan rộng toàn cầu từ tháng 9 đến tháng 11 năm 1918 và gây tử vong cao. Ở nhiều nước, đợt dịch thứ ba xảy ra vào đầu năm 1919 [21]. Biểu hiện lâm sàng tương tự nhau khiến các nhà quan sát thời đó kết luận ban đầu rằng họ đã nhìn thấy cùng một căn bệnh trong các đợt liên tiếp. Các dạng bệnh nhẹ ở cả 3 đợt là giống hệt nhau và là điển hình của bệnh cúm được ghi nhận trong đại dịch năm 1889 và trong những năm giữa các trận đại dịch trước đó. Nhìn lại, ngay cả những tiến triển nhanh chóng từ nhiễm cúm không biến chứng đến viêm phổi gây tử vong, có một dấu hiệu nhận biết của đợt dịch mùa thu và mùa đông năm 1918-1919, đã được ghi nhận trong một số trường hợp tương đối nghiêm trọng của đợt dịch mùa xuân. Do đó, sự khác biệt giữa các đợt dịch dường như chủ yếu ở tần suất xảy ra cao hơn nhiều của các trường hợp biến chứng, nặng và gây tử vong trong 2 đợt dịch sau.
Nhưng 3 đợt đại dịch cúm lan rộng trong vòng 1 năm, xảy ra liên tiếp nhanh chóng, chỉ với những khoảng thời gian yên lặng ngắn nhất giữa chúng, là chưa từng có. Sự xuất hiện, và ở một mức độ nào đó, của các đợt bùng phát tái diễn hàng năm, được thúc đẩy bởi sự trôi dạt của kháng nguyên virus, với một loại virus biến thể kháng nguyên mới để trở nên chiếm ưu thế sau mỗi 2-3 năm. Nếu không có sự trôi dạt như vậy, virus cúm lưu hành ở người có lẽ sẽ biến mất một khi khả năng miễn dịch của quần thể đã đạt đến ngưỡng tới hạn, là ngưỡng đủ giới hạn sự lây lan virus thêm nữa. Thời gian và khoảng cách của dịch cúm trong những năm giữa các đại dịch đã tạo điều kiện tích lũy trong nhiều thập kỷ. Các yếu tố được cho là chịu trách nhiệm bao gồm miễn dịch quần thể một phần hạn chế sự lây lan của virus trong mọi trường hợp, ngoại trừ những tình huống thuận lợi nhất, chẳng hạn nhiệt độ môi trường và nhiệt độ mũi người thấp hơn (có lợi cho các loài virus không chịu nhiệt như cúm), độ ẩm tối ưu, tình trạng đông đúc trong nhà gia tăng, thông khí không tốt do đóng cửa sổ và luồng không khí dưới mức tối ưu.
Tuy nhiên, các yếu tố như vậy không thể giải thích được 3 đợt đại dịch năm 1918- 1919, lần lượt xảy ra vào mùa xuân-hè, hè-thu và đông (của Bắc Bán cầu). Hai đợt đầu tiên xảy ra vào thời điểm trong năm thường không thuận lợi cho sự lây lan virus cúm. Đợt thứ hai gây bùng phát đồng thời ở Bắc và Nam Bán cầu từ tháng 9 đến tháng 11. Hơn nữa, các giai đoạn giữa những đợt dịch quá ngắn đến mức gần như không thể phát hiện được ở một số nơi. Giải thích dịch tễ học về sự sụt giảm mạnh các trường hợp ở đợt thứ nhất và thứ hai với sự gia tăng mạnh các trường hợp của đợt thứ hai và thứ ba là khó khăn. Giả sử ngay cả khả năng miễn dịch sau nhiễm bệnh thoáng qua, thì làm thế nào những người dễ mắc bệnh, là quá ít để có thể duy trì lây truyền ở 1 điểm, đủ để bắt đầu một đợt đại dịch bùng nổ mới vài tuần sau đó? Có phải virus đột biến sâu sắc và gần như đồng thời trên toàn thế giới, trong những giai đoạn ngắn giữa các đợt dịch liên tiếp? Có được sự trôi dạt virus đủ để tạo ra các chủng cúm mới có khả năng thoát khỏi tính miễn dịch quần thể được cho là mất nhiều năm lưu hành toàn cầu, chứ không phải vài tuần lưu hành tại chổ. Và nếu có xảy ra, những virus đột biến như vậy thường mất nhiều tháng để lây lan trên toàn thế giới.
Vào thời điểm bắt đầu các đại dịch cúm mùa khác, những đợt biệt lập liên tiếp trong vòng một năm chưa được báo cáo. Ví dụ, đại dịch năm 1889 bắt đầu vào cuối mùa xuân năm 1889 và mất vài tháng để lan rộng khắp thế giới, đỉnh điểm ở Bắc Âu và Hoa Kỳ vào cuối năm 1889 hoặc đầu năm 1890. Đỉnh tái phát thứ hai vào cuối mùa xuân năm 1891 (nhiều hơn hơn một năm sau khi xảy ra đại dịch đầu tiên) và lần thứ ba vào đầu năm 1892 [21]. Như đúng với đại dịch 1918, lần tái phát thứ hai năm 1891 đã gây ra nhiều tử vong nhất. Tuy nhiên, 3 lần tái phát vào năm 1889- 1892, trải dài trên 3 năm, trái ngược với 1918-1919, khi các đợt tiếp theo được thấy ở các quốc gia riêng lẻ thường tập trung vào ̴ tháng 8-9.
Điều gì đã mang lại cho virus 1918 khả năng chưa từng có để gây ra các đợt đại dịch liên tiếp nhanh chóng vẫn chưa rõ ràng. Bởi vì các mẫu virus đại dịch 1918 mà chúng tôi đã xác định được là từ những bệnh nhân của đợt thứ hai [16], nên không gì có thể khẳng định cùng một loại virus hoặc các biến thể của nó lưu hành trong cả đợt thứ nhất và thứ ba. Dữ liệu từ năm 1918 cho thấy rằng những người bị nhiễm trong đợt thứ hai có thể đã được bảo vệ khỏi bệnh cúm ở đợt thứ ba. Nhưng một số ít dữ liệu về khả năng bảo vệ trong đợt thứ hai và thứ ba sau khi bị nhiễm ở đợt thứ nhất là không thuyết phục và không giải quyết được câu hỏi liệu đợt đầu tiên có phải do cùng một loại virus hay không, liệu các sự kiện tiến hóa di truyền lớn có xảy ra ngay cả khi đại dịch đang bùng nổ và tiến triển. Chỉ các mẫu xét nghiệm ở người dương tính với RNA cúm từ trước 1918 và từ cả 3 đợt, mới có thể trả lời câu hỏi này.
Nguồn gốc vật chủ của virus đại dịch là gì?
Dữ liệu về trình tự gen virus hiện nay cho thấy toàn bộ virus 1918 là một chủng mới xuất hiện đối với con người, hoặc xuất hiện ngay trước 1918, do đó, chúng không phải là một loại virus tái tổ hợp được tạo ra từ các chủng cũ để hình thành 1 hoặc nhiều gen mới, chẳng hạn như những chủng gây ra đại dịch 1957 và 1968. Ngược lại, virus 1918 có vẻ như là một loại virus gia cầm có nguồn gốc chưa xác định [17,19], vì 8 đoạn gen của nó có sự khác biệt đáng kể so với gen cúm gia cầm vào thời điểm đó. Trình tự gen virus cúm từ một số mẫu của loài chim hoang dã được thu thập khoảng năm 1918 cho thấy có một ít khác biệt với các virus gia cầm được phân lập ngày nay, chỉ ra rằng virus gia cầm có khả năng biến đổi kháng nguyên một chút bên trong vật chủ tự nhiên của chúng trong một khoảng thời gian dài [24,25].
Ví dụ, trình tự gen nucleoprotein (NP) 1918 tương tự như virus tìm thấy ở chim hoang dã ở cấp độ sắp xếp chuỗi acid amin nhưng rất khác nhau ở cấp độ sắp xếp chuỗi nucleotide, cho thấy khoảng cách tiến hóa đáng kể giữa nguồn gốc của NP 1918 và trình tự gen NP hiện tại trong các chủng chim hoang dã. Một cách để xem xét khoảng cách tiến hóa của gen là so sánh các tỷ lệ thay thế nucleotide đồng nghĩa cho nucleotide không đồng nghĩa. Một thay thế đồng nghĩa biểu hiện một thay đổi im lặng, thay đổi nucleotide trong một codon mà nó không dẫn đến thay thế acid amin. Một thay thế nucleotide không đồng nghĩa là thay đổi nucleotide trong một codon mà nó dẫn đến sự thay thế acid amin. Nói chung, một gen virus chịu áp lực đào thải của miễn dịch hoặc thích nghi với vật chủ mới cho ta biết rằng tỷ lệ thay thế nucleotide không đồng nghĩa nhiều hơn, trong khi virus dưới áp lực chọn lọc nhỏ thì tích lũy thay đổi chủ yếu là thay đổi đồng nghĩa. Vì ít hoặc không có áp lực chọn lọc nào tác động lên những thay đổi đồng nghĩa, chúng được cho là phản ánh khoảng cách tiến hóa.
Do các phân đoạn gen năm 1918 có nhiều thay thế đồng nghĩa từ các trình tự đã biết của các chủng chim hoang dã hơn dự kiến, nên chúng dường như không phải xuất hiện trực tiếp từ một loại virus cúm gia cầm tương tự với những chủng đã được giải trình tự cho đến nay. Điều này đặc biệt rõ ràng khi người ta kiểm tra sự khác biệt ở các codon giáng hóa 4 lần, có các thay đổi đồng nghĩa, ở vị trí codon thứ ba, bất kỳ nucleotide nào trong số 4 nucleotide đều có thể được thay thế mà không làm thay đổi việc tạo ra acid amin. Đồng thời, trình tự gen 1918 có quá ít sự khác biệt về acid amin so với các chủng virus chim hoang dã đã mất nhiều năm thích nghi chỉ ở vật chủ trung gian là người hoặc lợn. Một lời giải thích có khả năng là những đoạn gen bất thường này bắt nguồn từ một ổ virus cúm chưa từng được xác định hoặc lấy mẫu. Tất cả những phát hiện này đặt ra câu hỏi: virus 1918 đã bắt nguồn từ đâu?
Trái với cấu trúc di truyền của virus đại dịch 1918, các phân đoạn gen mới của virus đại dịch 1957 và 1968 được tái tổ hợp đều có nguồn gốc từ virus cúm gia cầm Á-Âu [26]; cả hai loại virus ở người phát sinh theo cùng một cơ chế tái tổ hợp từ một chủng thủy cầm hoang dã Á-Âu với chủng H1N1 ở người lưu hành trước đó. Chứng minh giả thuyết rằng virus gây ra đại dịch 1918 có nguồn gốc khác biệt rõ rệt đòi hỏi các mẫu cúm ở người lưu hành trước năm 1918 và các mẫu của chủng cúm ở chim gần giống hơn với trình tự 1918.
Cơ sở sinh học của mầm bệnh virus đại dịch 1918 là gì?
Phân tích giải trình tự gen đơn độc không cung cấp thông tin về khả năng gây bệnh của virus 1918. Một loạt các thí nghiệm đang được tiến hành để mô hình hóa độc lực virus trong ống nghiệm và trên các mô hình động vật bằng cách sử dụng các cấu trúc virus chứa gen 1918 được tạo ra bởi sao chép ngược.
Virus cúm đòi hỏi phải gắn protein ngưng kết hồng cầu (HA) với các thụ thể của acid sialic trên bề mặt tế bào chủ. Receptor liên kết với thụ thể HA của virus để thích nghi để lây cho chim và thích nghi để lây cho người là khác nhau. Các chủng virus cúm thích nghi với các loài chim ưu tiên liên kết các thụ thể acid sialic với các liên kết đường α (2-3) [27-29]. Virus cúm thích nghi ở người được cho là ưu tiên liên kết các thụ thể với các liên kết đường α (2-6). Việc chuyển đổi cấu trúc receptor cúm gia cầm này đòi hỏi virus chỉ có 1 thay đổi acid amin và HA của cả 5 loại virus 1918 được giải trình tự đều có sự thay đổi này, điều này cho thấy đây có thể là một bước quan trọng trong việc thích nghi của virus với vật chủ con người. Sự thay đổi acid amin thứ hai cũng có thể làm gia tăng đáng kể sự liên kết của virus với thụ thể ở người, nhưng chỉ có 3 trong số 5 chuỗi HA 1918 có sự thay đổi này [16].
Điều này có nghĩa là ít nhất 2 biến thể gắn kết với receptor H1N1 được tạo ra ở 1918: 1 biến thể có gắn kết ái lực cao với thụ thể người và 1 biến thể với gắn kết ái lực hỗn hợp với cả thụ thể của gia cầm và người. Không có dấu hiệu nào về địa lý hay thời gian sống minh chứng cho việc một trong những biến thể này là nguồn gốc của biến thể kia, cũng không có sự khác biệt nhất quán giữa bệnh sử hay đặc điểm mô bệnh học của 5 bệnh nhân bị nhiễm chúng. Cho dù virus có khả năng lây truyền như nhau vào năm 1918, nhưng liệu chúng có mô hình sao chép giống hệt nhau ở đường hô hấp hay không và liệu một hay tất cả đều lưu hành trong đợt đại dịch thứ nhất và thứ ba, vẫn chưa được biết rõ.
Trong một loạt các thí nghiệm trên cơ thể sống, những virus cúm tái tổ hợp có chứa từ 1 đến 5 đoạn gen của virus 1918 đã được tạo ra. Những cấu trúc mang HA và NA 1918 đều có khả năng gây bệnh cao ở chuột [31]. Hơn nữa, vi phân tích về sự sắp xếp gen được thực hiện trên toàn bộ mô phổi của chuột bị nhiễm HA/NA 1918 tái tổ hợp cho thấy sự gia tăng điều hòa tăng biểu hiện các gen liên quan đến chết tế bào theo chu trình, tổn thương mô và tổn thương oxy hóa [32]. Những phát hiện này thực sự đáng ngạc nhiên bởi các virus có gen 1918 không thích nghi được với chuột; các thí nghiệm được kiểm soát có chuột bị nhiễm virus người cho thấy ít gây bệnh và hạn chế sự nhân lên của virus. Phổi của động vật bị nhiễm cấu trúc HA/NA 1918 cho thấy hoại tử biểu mô phế quản và phế nang cũng như thâm nhiễm viêm rõ rệt, cho thấy rằng HA 1918 (và có thể cả NA) có chứa các yếu tố độc lực đối với chuột. Cơ sở di truyền kiểu gen virus gây bệnh này chưa được lập bản đồ gen. Liệu mầm bệnh ở chuột có cho ta hình dung được mô hình khả năng gây bệnh ở người hiệu quả hay không vẫn chưa rõ ràng. Vai trò tiềm năng của các protein 1918 khác, khi đơn lẻ và khi kết hợp, cũng chưa được biết. Các thí nghiệm để xác định cơ sở di truyền của độc lực virus 1918 trong các mô hình động vật khác nhau đã được lên kế hoạch. Những thí nghiệm này có thể giúp định dạng thành phần virus cùng khả năng gây bệnh bất thường của virus 1918 nhưng không thể giải quyết được vấn đề rằng liệu các yếu tố vật chủ cụ thể năm 1918 có giải thích mô hình tỷ lệ tử vong độc đáo do cúm hay không.
Tại sao Virus 1918 có thể giết chết nhiều người trưởng thành khỏe mạnh?
Đường cong tử vong do cúm theo tuổi từ trước đến nay trong ít nhất 150 năm qua, có dạng hình chữ U (Hình 2), ta thấy mức tử vong cao nhất ở trẻ nhỏ và người già, với tần suất tử vong tương đối thấp ở mọi lứa tuổi ở giữa. Ngược lại, tỷ lệ tử vong theo độ tuổi cụ thể trong đại dịch năm 1918 thể hiện một dạng biểu đồ riêng biệt chưa từng được ghi nhận từ trước đến nay: một đường cong hình chữ W, tương tự như đường cong hình chữ U quen thuộc nhưng có thêm một phần ba (giữa ) có đỉnh tử vong rõ rệt ở thanh niên trong khoảng 20-40 tuổi. Chẳng hạn, tỷ lệ tử vong do cúm và viêm phổi đối với những người 15-34 tuổi trong khoảng năm 1918-1919, cao hơn >20 lần so với những năm trước [35]. Nhìn chung, gần một nửa số ca tử vong liên quan đến cúm trong đại dịch 1918 là ở người trưởng thành 20-40 tuổi, bất thường duy nhất trong năm đại dịch đó. Đại dịch 1918 cũng là đại dịch duy nhất trong số các đại dịch cúm có nguy cơ tử vong hoàn toàn do cúm cao hơn ở những người <65 tuổi so với những người >65; những người <65 tuổi chiếm >99% tổng số ca tử vong liên quan đến cúm khoảng năm 1918-1919. So với nhóm <65 tuổi chiếm 36% tổng số ca tử vong liên quan đến cúm trong đại dịch H2N2 năm 1957 và 48% trong đại dịch H3N2 năm 1968 [33].
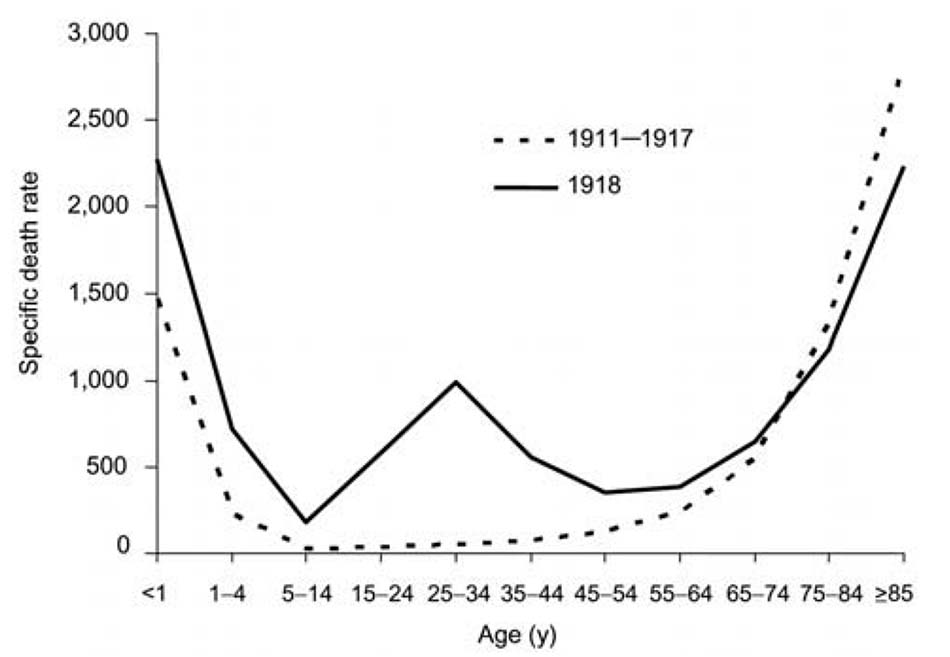
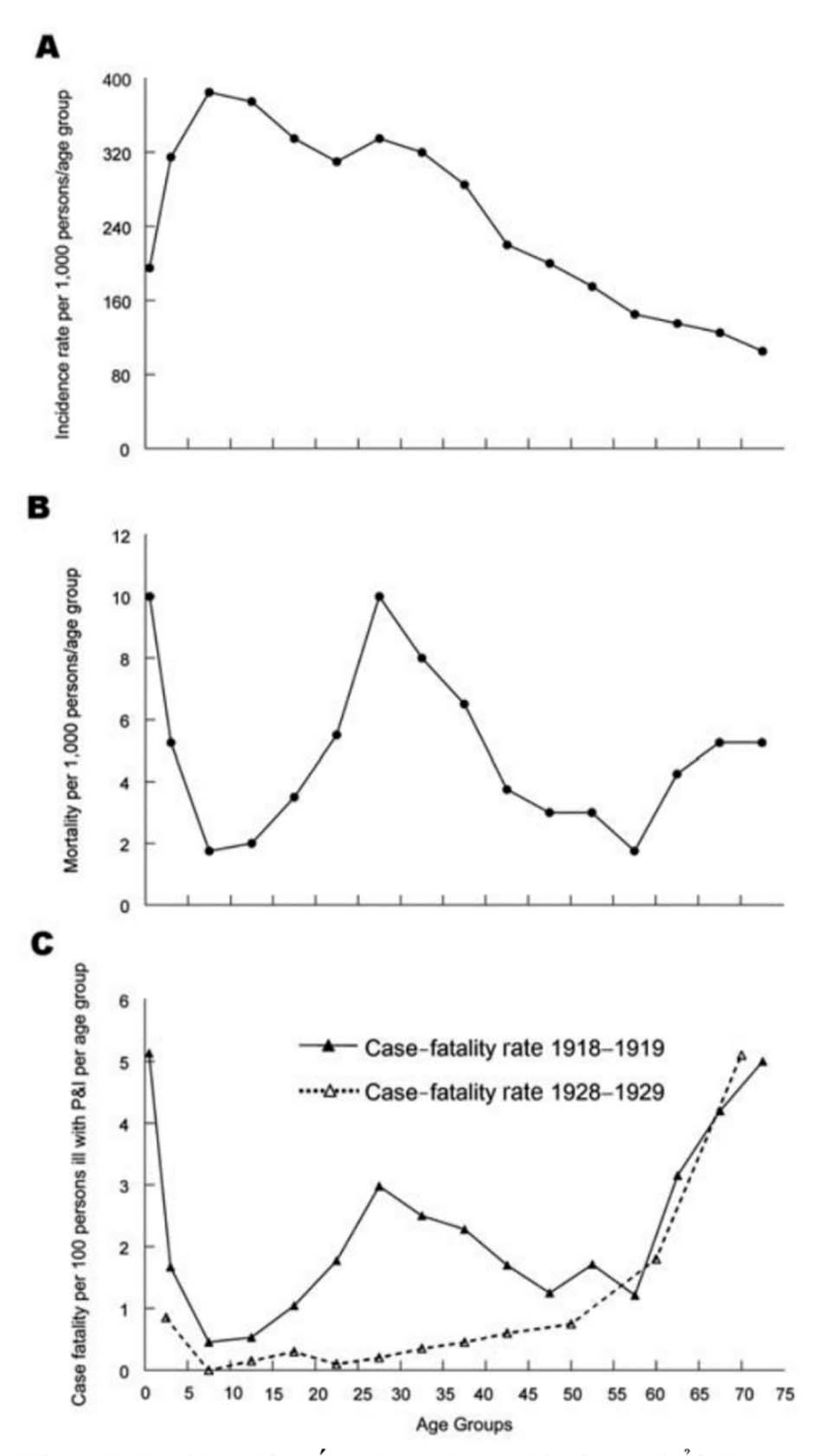
Một góc nhìn sắc nét hơn xuất hiện khi năm 1918 tỷ lệ mắc cúm theo tuổi cụ thể [21] được sử dụng để hiệu chỉnh đường cong tử vong dạng W (Hình 3, A, B, và C). Những người <35 tuổi vào năm 1918 có tỷ lệ mắc cúm cao không tương xứng (Hình 3, bảng A).
Nhưng ngay cả sau khi điều chỉnh các trường hợp tử vong/độ tuổi cụ thể theo tỷ lệ bị nhiễm trên lâm sàng theo độ tuổi cụ thể (Hình 3, bảng B), đường cong hình chữ W với đỉnh tử vong ở trường hợp những người trẻ tuổi vẫn còn và có sự khác biệt đáng kể so với đường cong tử vong theo tuổi cụ thể hình chữ U thường thấy ở cúm trong các năm khác, ví dụ 1928- 1929 (Hình 3, bảng C). Ngoài ra, vào năm 1918, những người từ 5 đến 14 tuổi chiếm số ca cúm không tương xứng, nhưng có tỷ lệ tử vong và viêm phổi do cúm thấp hơn nhiều so với các nhóm tuổi khác. Để giải thích biểu đồ này, chúng ta phải nhìn xa hơn về các đặc tính của virus đối với vật chủ và các yếu tố môi trường, có thể bao gồm cả miễn dịch học (ví dụ, tăng cường nhiễm bệnh phụ thuộc kháng thể liên quan đến phơi nhiễm virus trước đó [38]) và tiếp xúc với các đồng yếu tố nguy cơ chẳng hạn như những tác nhân đồng nhiễm, thuốc và yếu tố môi trường.
Một giả thuyết có thể giải thích một phần những điều này là tự virus 1918 có độc lực cao, chỉ được trung hòa ở những bệnh nhân sinh trước năm 1889, ví dụ, do quá trình tiếp xúc với virus lưu hành sau đó có khả năng miễn dịch bảo vệ chống lại một phần chủng virus 1918 chỉ có ở những người đủ độ tuổi (> 35 tuổi) đã từng bị nhiễm trong thời kỳ trước đó [35]. Nhưng lý thuyết này sẽ đưa ra một nghịch lý khác kèm theo: một loại virus tiền thân tiềm ẩn không phát hiện được dấu vết cho đến ngày nay, đã phải xuất hiện và biến mất trước năm 1889, và sau đó xuất hiện trở lại sau hơn 3 thập kỷ.
Liệu một đại dịch giống như 1918 có thể xuất hiện trở lại?
Nếu vậy, chúng ta có thể làm gì với nó? Trong diễn biến bệnh và các đặc điểm bệnh lý của nó, đại dịch 1918 so với các đại dịch trước và sau đó, có sự khác nhau về mức độ, nhưng không khác nhau về loại dịch. Mặc dù số lượng tử vong toàn cầu rất cao, nhưng hầu hết các trường hợp cúm vào năm 1918 (>95% ở hầu hết các địa phương thuộc những quốc gia công nghiệp hóa) đều nhẹ và không thể phân biệt với các trường hợp cúm ngày nay. Hơn nữa, các thí nghiệm với virus cúm tái tổ hợp có chứa gen từ virus 1918 cho thấy các virus giống như virus 1918 và tương tự 1918 sẽ nhạy cảm như các chủng virus điển hình khác đối với các loại thuốc chống virus rimantadine và oseltamivir đã được FDA phê duyệt.
Tuy nhiên, có một số đặc điểm chỉ có ở đại dịch năm 1918: đáng chú ý nhất là tỷ lệ tử vong cao gấp 5-20 lần so với dự kiến. Về mặt lâm sàng và bệnh học, tỷ lệ tử vong cao này có vẻ là kết quả của nhiều yếu tố, bao gồm tỷ lệ cao của nhiễm khuẩn đường hô hấp nặng nề và phức tạp, chứ không phải là do các hệ cơ quan khác bị ảnh hưởng bởi virus cúm. Ngoài ra, số người tử vong tập trung một cách bất thường ở nhóm tuổi trẻ. Cuối cùng, vào năm 1918, 3 đợt cúm tái phát riêng biệt xảy ra với tốc độ nhanh bất thường, dẫn đến 3 đợt đại dịch bùng nổ trong vòng một năm (Hình 1). Mỗi đặc điểm độc đáo này có thể phản ánh các đặc điểm di truyền của virus 1918, nhưng để hiểu biết rõ về chúng cũng cần phải kiểm tra các yếu tố môi trường và vật chủ.
Cho đến khi chúng ta có thể xác định được yếu tố nào trong số những yếu tố này đã đưa đến mô hình tử vong được quan sát thấy và tìm hiểu thêm về sự hình thành của đại dịch, thì việc dự đoán chỉ là mô phỏng lý thuyết. Chúng ta chỉ có thể kết luận rằng vì nó đã xảy ra một lần, các điều kiện tương tự có thể dẫn đến xảy ra một đại dịch tàn khốc không kém.
Giống như virus 1918, H5N1 là virus gia cầm [39], cũng là một chủng virus không mấy liên quan. Con đường tiến hóa dẫn đến sự xuất hiện của đại dịch năm 1918 hoàn toàn không được biết, nhưng dường như nó khác biệt ở nhiều khía cạnh so với dịch H5N1 ở hiện tại. Không có dữ liệu lịch sử nào trước đây, vào năm 1918 hoặc trong bất kỳ đại dịch nào khác, để xác định rằng một loại virus tiền thân của đại dịch gây ra đợt bùng phát bệnh cao ở gia cầm và không có virus cúm gia cầm gây bệnh cao (HPAI), bao gồm H5N1 và một số chủng khác, đã từng được biết là gây ra một dịch bệnh lớn ở người, chứ chưa nói đến một đại dịch. Trong khi dữ liệu về khả năng thích nghi tế bào người của virus cúm đang bắt đầu được hiểu ở cấp độ phân tử (ví dụ: gắn kết thụ thể), thì cơ sở cho sự thích nghi của virus để lây lan từ người sang người một cách hiệu quả, điều kiện tiên quyết chính cho sự xuất hiện đại dịch, vẫn chưa được biết đối với bất kỳ virus cúm nào. Virus 1918 có được đặc điểm này, nhưng chúng tôi không biết làm thế nào và hiện tại chúng tôi không có cách nào để biết liệu virus H5N1 đang trong quá trình tiến tới có thể truyền bệnh từ người sang người hay không. Mặc dù có sự bùng nổ về dữ liệu về virus 1918 trong thập kỷ qua, nhưng chúng ta không thể hiểu rõ hơn về sự xuất hiện của đại dịch năm 2006 so với những gì chúng ta hiểu về nguy cơ xuất hiện cúm lợn H1N1 năm 1976.
Ngay cả với các loại thuốc chống virus và kháng khuẩn hiện đại, vaccine và các kiến thức phòng ngừa, sự trở lại của một loại virus gây bệnh tương tự khả năng gây bệnh của virus 1918 có thể sẽ giết chết hơn 100 triệu người trên toàn thế giới. Một loại virus đại dịch với tiềm năng gây bệnh (được cho là) của một số đợt bùng phát H5N1 gần đây có thể gây ra tử vong nhiều đáng kể.
Cho dù do yếu tố virus, vật chủ hay môi trường, thì virus 1918 gây ra đợt đầu tiên hoặc “mùa xuân” không liên quan đến khả năng gây bệnh đặc biệt của đợt thứ hai (mùa thu) và thứ ba (mùa đông). Xác định một trường hợp dương tính với RNA cúm từ đợt đầu tiên có thể chỉ ra cơ sở di truyền cho độc lực virus bằng cách chú ý đến sự khác biệt trong trình tự gen virus. Việc xác định các mẫu RNA cúm ở người trước năm 1918 sẽ giúp chúng ta hiểu được thời điểm xuất hiện của virus 1918. Giám sát theo dõi và giải trình tự bộ gen của một số lượng lớn virus cúm động vật sẽ giúp chúng ta hiểu được cơ sở di truyền của sự thích nghi vật chủ và mức độ của ổ chứa virus cúm trong tự nhiên. Hiểu về đại dịch cúm nói chung đòi hỏi phải hiểu về đại dịch năm 1918 ở tất cả các khía cạnh lịch sử, dịch tễ và sinh học của nó.
Tài liệu tham khảo












{kind=link}